I suppose a more important question is why we neantherthals did not
start testing those hypotheses sooner? Murray apparently began to,
but
gait labs were fewer and further between in those days, and the
process of kinematic recording was more tedious.
I am at a loss to contribute an original answer to the question at
hand ("What got us walking on two legs?"), but if someone does come
up
with a nifty idea, we should be readier to pounce on it with
experimental corroboration or refutation, rather than let it harden
into another set of dogmatic stone tablets.
I hate to see Inman and Saunders discredited for having come up with
a
"wrong" idea. The ingenuity of their invention is remarkable. The
determinants of gait are simply another of many bright signs of their
hard-working and productive minds that were light years ahead of most
others' in the kinesiology of half a century ago. We should instead
chide ourselves for having regarded the "major determinants" as
sacrosanct.
There was an excellent paper at the recent ISB meeting in Calgary
that
I believe comes the closest I've seen to testing a theory of human
gait. It was by Marcus Pandy and his PhD student Anderson (Marcus
presented the paper although it is his student's work I believe) from
the University of Texas at Austin. The paper was entitled "A dynamic
optimization solution for one complete cycle of human gait".
Many
muscles were modelled and the objective function minimised was the
energy output (chemical not mechanical) of the muscles. The system
predicted muscle forces (the phasing of which agreed well with EMG),
ground reaction forces (which agreed well with what your good old
Kistler would give you but the system knew nothing a priori about the
double bump shape, etc.), joint torques, joint kinematics, etc.
The 3D graphical output was really quite stunning and I was convinced
that the authors were really onto something special (by the way the
model also took 50 hours of supercomputer time to run!). A second
paper they presented, which showed that static optimisation (i.e. where
time is not explicit in the objective function) performs as well as
their approach (but is far less computationally demanding), took a
little of the shine off their first paper. Bottom line, however, is
that this was without doubt the most sophisticated simulation I have
seen of human gait and it will enable us to explore this "unified
theory of bipedal gait" in more depth.
Kit Vaughan
University of Cape Town
How the human came to evolve to locomote in a bipedal fashion versus
a
quadrupedal fashion is certainly an interesting question. Another
very
separate question is why does each human individual currently walk
the way
they do? In other words, why do we choose the specific
gait pattern which
characterizes the way we walk?
My view is that each individual is driven to walk in their own characteristic
way for three main purposes:
1) to move from point A to point B in the most efficient manner (i.e. least metabolic energy expended) given their specific morphologic characteristics and functional capacity, and
2) to move from point A to point B in the least painful manner (i.e. pain avoidance), and
3) to move from point A to point B to avoid injury to their body (i.e. injury avoidance).
I am sure that these are not earth-shattering principals to anyone on this list, however, in my experience primarily as a clinician, and secondly as a researcher and educator, these three purposes of gait seem to be upheld on a daily basis in my practice.
The determinants of gait model proposed by Saunders, et al in
1953, is still
an excellent way to teach students how an animal, such as a human,
may have
chosen to walk the way it does. I strongly believe that it still
has
application in the clinical setting for those involved in the diagnosis
and
treatment of gait abnormalities since it greatly simplifies a very
complex
subject which is easily understood by the student by using a unifying
principle: that humans move the way they do during walking
to minimize
energy expenditure by minimizing the vertical and/or horizontal
oscillations
of the center of mass.
In regard to foot and lower extremity pathology, we often see
individuals in
the clinical setting with minor painful abnormalities of their foot
who
change their gait patterns for days, months and even years to avoid
painful
conditions in their lower extremities. In addition, injury avoidance
may
cause alterations in gait patterns. For example, patients with
certain foot
types which predispose them to chronic lateral ankle instability may
habitually walk with a shorter stride and wider base of gait than normal
to
avoid inversion ankle sprains.
The complexity of why we walk the way we do is a fascinating subject
and I
appreciate the opportunity to share my thoughts with such an interested
and
educated group of individuals.
Cheers,
Kevin A. Kirby, D.P.M.
Assistant Clinical Professor of Biomechanics
California College of Podiatric Medicine
Private Practice:
2626 N Street
Sacramento, CA 95816
Voice: (916) 456-4768 Fax: (916) 451-6014
Ref: Tucker CA, Ramirez J, Krebs DE, Riley PO. Center of gravity dynamic
stability in normal and vestibulopathic gait. Gait & Posture, 1998;8:117-123
David E. Krebs, PhD, PT
Professor and Director, MGH Biomotion Laboratory
MGH Institute of Health Professions
101 Merrimac Street
Boston, MA 02114-4719
V: 617 726 8016
1. Gard, SA and Childress, DS, "The effect of pelvic list on the vertical
displacement of the trunk during normal walking",
Gait and Posture 5 (1997), 223-338.
2. Gard, SA and Childress, DS, "The Influence of Stance-Phase Knee Flesion
on the Vertical Displacement of Trunk
During Normal Walking", Archives of Physical Medicine and Rehabilitation,
Vol. 80, pp. 26-32 (Jan. 1999).
3. Gard, SA, Knox, EH, and Childress, DS, "Two-Dimensional Representation
of Three-Dimensional Pelvic Motion
During Human Walking: An Example of How Projections Can Be Misleading",
J. Biomechanics, Vol. 29, No. 10, pp.
1387-1391.
4. Childress, DS, and Gard, SA, "Vertical Movement of the Trunk in Human
Walking", Abstracts, 17th ISB Congress, p.
116, Calgary, Aug. 8-13, 1999.
We are sending you copies of these papers along with a copy of the original
paper by Saunders, Inman, and Eberhart,
(1953).
The first two papers (publications 1 & 2) deal with pelvic obliquity
and knee flexion, which we believe act importantly as shock absorbers for
the body. Consequently, their primary action must come at the time of heel
contact, not later during midstance. It's a matter of timing. By the time
of midstance, or just thereafter, the action of the knee and the action
of thepelvis have been completed. Inman's own data supports this observation.
The role of these two movements can't be to
reduce vertical movement of the trunk at mid-stance. Our data demonstrate
this. Pelvic obliquity and knee flexion are
important attributes of gait but we think their influence is primarily
to reduce shock at the beginning of double support, not
to reduce vertical movement of the trunk (e.g. supress the height at
midstance). They could do both, but our papers
indicate otherwise.
We did not set out to try to show that the second and third determinants
were questionable. We were interested in toe
clearance during swing through. Steve Gard was doing an investigation
of toe clearance as part of his Ph.D. work.
Clearance is a particularly important aspect of an amputee's gait.
Our feeling was that if pelvic obliquity and knee flexion
lowered the COM near midstance that the human walker would have to
compensate for the lowering of the trunk with
flexion of the knee and/or dorsiflexion of the foot to achieve foot
clearance during swing-through. Careful examination
showed that the trunk was not lowered at midstance in the way Saunders
et al. had suggested. I held on to the 2nd and
3rd determinants (having believed them for 25 years) until the data
and Steve convinced me otherwise. If you read the
original paper by Saunders, Inman, and Eberhart carefully, you find
that they based their ideas on "thought experiments".
No data was presented. Thought experiments are OK but ultimately they
need to be backed up with data.
I believe the six determinants described are really six attributes (traits,
characteristics, etc.) of gait. The first determinant is
pelvic rotation. Pelvic rotation enables a person to take a longer
step, almost like lengthening the legs. The purpose of
pelvic rotation seems mainly a way to lengthen the stride, Whatever
energy might be saved in vertical excursion would
more than be made up for by muscular energy required for the trunk
rotation.
The second determinant is pelvic (tilt) obliquity and the third is stance
phase knee flexion. Both have already been dealt
with in our papers and above.
The fourth and fifth determinants are described as foot/knee mechanisms.
There is a lot of hand waving about these two
determinants by Saunders, et al. and they are not precisely separated
in the original article, although they are supposed to
smooth out the pathway to the COM. (We think ankle motion is important;
again, for shock absorption.) By the time
Saunders et al. get to the "Summary" of their paper, they only list
five mechanisms for the six determinants and have
changed the third determinant from "Knee Flexion in Stance Phase" to
"Knee and Hip Flexion". Interpreters of the six
determinants have separated them in several different ways. For example,
Whittle, in his book, "Gait Analysis" makes the
4th determinant "Ankle Mechanism" and the 5th, "Foot Mechanism". McMahon,
in his book, "Muscles, Reflexes, and
Locomotion", goes so far as to make "Compass Gait" the 1st determinant.
He then moves the others down in order,
leaving out the original fourth determinant and making the 5th determinant,
"Plantar flexion of the stance ankle." Actually, I
could accept "compass gait" as a possible determinant.
The 6th determinant is "Lateral displacement of the pelvis". I leave
interpretation of this one to you. Personally, I don't buy
it as any great saver of energy. It's more a trait, attribute, or characteristic
of human gait than a determinant.
Saunders, et al. show the intersection of horizontal and vertical displacements
on page 546 of their original paper. In their
book, "Human Walking", Figure 1.1 shows a butterfly-shaped trace for
this intesection. It actually should be a "U"-shaped
curve. We're fairly certain they drew the butterfly shape because that
is what you typically get when you record from a
marker near the COM on the back or front of the body during walking.
We believe the butterfly is a consequence of the
added movement of pelvic rotation and the marker not being at the point
of rotation. We show why this is probably so in
publication 3, which appeared in the J. of Biomechanics. By the way,
Marey in his book, "Animal Mechanism: Terrestial
and Aerial Locomotion", p. 119, (1893) draws this curve correctly,
we think, in the shape of a "U".
Please know that I am not anti-Inman. I knew him in his later years
and have the greatest respect for what he and the
others did at Berkeley and in San Francisco after the war. What they
accomplished in one or two years of work is
amazing. I refer to Inman's book often and recommend it to my students.
I just can't buy the concept of energy
conservation in the six traits as proposed. As a matter of fact, I
doubt that energy conservation is the primary goal in
human walking. I've yet to see a single performance index determine
human action.
Publication 4, is an introduction to a paper that is now in progress.
Out of this work we describe all of the gait
parameters mathematically in terms of lengths and constants. I presented
an advanced version at ISB.
Dudley S. Childress, Ph.D.
Prof. of PM&R and Biomedical Engineering, Northwestern University
345 East Superior St., Chicago, IL 60611 USA, Rm. 1441
TEL: 312.908-8560, FAX: 312.908.6526
Thanks again to Chris for stirring the pot!
As Chris noted, we had an extensive discussion some months ago of Saunders, Inman, & Eberhart's "The Major Determinants of Normal and Pathological Gait." Now we seem to be starting such a discussion again. I would hate to see the discussion of the origins of alternating bipedal locomotion be derailed by our getting bogged down in the "Determinants" again, but the paper had, I believe, a fundamental deficiency that must be considered before we can go beyond it.
That deficiency was that the paper focussed on mechanisms for minimizing the magnitudes of vertical and lateral oscillations of the center of mass, but made no mention of the fact that, in the direction of progression, the center of mass MUST speed up and slow down with every step. This is an inevitable consequence of the fact that in alternating bipedal walking, the center of support is only directly under the center of mass at four points in each gait cycle - once in the middle of each double support phase, and once in the middle of each single support phase.
The rest of the time, the point of support is either in front of the center of mass, causing a deceleration of forward progression, or behind the center of mass, causing an acceleration of forward progression. Thus, the progressional velocity of the body MUST speed up, and then slow down again, twice in every gait cycle. Consequently the kinetic energy of the body as a whole MUST increase and decrease in a cyclic manner with every step.
Now, in contrast to this unavoidable cyclic change in kinetic energy, the potential energy of the body, as represented by the vertical elevation of the body, is not required, by any fundamental principle that I know of, to change with every cycle. The "determinants" that Saunders, Inman, and Eberhart describe would be quite sufficient to allow the vertical oscillations of the body to be brought to zero, if such were desirable.
Because we never observe people walking with zero vertical oscillation, however, the only conclusion I can come to is that reducing the vertical oscillations of the body to zero is not desirable, in an energetic sense. That should not come as a surprise, because if the kinetic energy MUST change with every cycle, then there is an energetic advantage to having the potential energy of the body change in a complementary fashion, keeping the total mechanical energy of the body more uniform.
This concept is familiar to us in the simple pendulum, where total energy, or the sum of kinetic and potential energy, is constant, but the ratio of kinetic to potential energy continually changes in a cyclic manner.
I submit that this physical necessity of speeding up and slowing down with every step is a first principle in the energetics of walking, and that all the "determinants" are secondary responses. I think this explains why the "determinants" have not provided a basis for a comprehensive understanding of the energetics of human walking.
Paul Andrews wrote: "I hate to see Inman and Saunders discredited for having come up with a "wrong" idea. The ingenuity of their invention is remarkable. The determinants of gait are simply another of many bright signs of their hard-working and productive minds that were light years ahead of most others' in the kinesiology of half a century ago. We should instead chide ourselves for having regarded the "major determinants" as sacrosanct."
I agree whole-heartedly. One of the great privileges of my professional life was to work for almost 20 years with Inman and Eberhart. They were the prototype of an effective Orthopaedist/Engineer team and were extraordinarily creative and productive. Criticism is the essence of scientific progress, and need not imply contempt.
Kevin Kirby wrote: "... humans move the way they do during walking to minimize energy expenditure by minimizing the vertical and/or horizontal oscillations of the center of mass."
That was the message of the "determinants" paper. I think that message is misleading, for the reasons stated above. I think the "determinants" paper has actually hindered our understanding of the energetics of walking by distracting us from the "first principle" that the body center of mass MUST speed up and slow down with every step. Perhaps we should be paying a lot more attention to the nature of cyclic changes in each subject's progressional velocity.
Thanks to all for a splendid discussion!
Larry Lamoreux, Ph.D., Gait Dimension, Lafayette, CA, USA
Carl Sagan said in his book - The dragons of eden : speculations on
the
evolution of human intelligence, the ability to walk upright
on two legs
has played a key role in human life-style for more than 3 million
years.
Could there be a concurrent development and interdependence of bipedal
walking and intelligence during our evolution?
TSANG Tak Lung
Final Year Project student
The Hong Kong Polytechnic University
Great converstation: Larry Lamoreux previously wrote, the center
of mass
MUST speed up and slow down, and reducing the vertical oscillations
fo the
body to zero is not desireable. Therefore, this physical
necessity of
speeding up and slowing down is a first principle in the energetics
of
walking.
He also noted that the center of support is only under the center
of mass
twice/side during the cycle. This is attributed to the
need of the CoM to
initially be behind and then ahead of the center of support.
Chris then wrote of an assumption of the foot being like a wheel, but
then
offered a withdrawl of this concept based on Larry's submission.
I would like to stir the pot a bit by suggesting that Chris' assumption
of the
wheel is correct and that the center of support is not
any fixed point but
rather a mobile one, and that combining these two concepts will lead
to
further understanding of the origins of bipedal gait.
In the original Determinants of Gait, the foot is viewed as a rigid
body.
While this may make mathemathical modeling feasible, the foot has so
many
joints that considering it a rigid structure misses the elegance of
its
design as well as its actual functional contribution to the entire
structure
above it.
Consistant with Dr. Lamoreux description, if the CoM moves up and over
the
center of support, the support structure MUST permit the appropriate
motion
which allows for this to occur. The foot, therefore, must function
as a
sagittal plane pivot. Perry has described the three rocker
system of the
foot/ankle which precisely permits this function. It consists
of the of the
round underside of the heel, the dorsiflexing ankle and dorsiflexing
1st MTP
joint. Since each aspect move in sequence, no single center of
support
exists, but instead a transient "axis of rotation". While the
heel and ankle
are in close proximity, the MTP joint is quite a distance from the
other two.
It would therefore fit Chris' "wheel model" analogy, ie a series
of ground
contact points occuring sequentially. (In one of the most pathomechanic
of
feet, the rocker bottom displays this principle rather clearly.
The loss of
the normal pivotal rockers causes the foot to assume a shape consistent
with
its functional requirement. This is a clear visual of Wolfe's
Law, Form
Follows Function.)
As the foot transitioned from an arborial to terrestrial structure,
the
adaphistic great toe which would use plantarflexion for tree limb grasping
needed to evolve to a dorsiflexing great toe which would function as
the
final sagittal pivot. OJ Lewis published a series in the Journal
of Anatomy
in 1979 which showed how it was a rotational alignment change in the
lateral
column of the foot (inversion) which actually repositioned the peroneal
axis
of pull on the 1st ray and thus permitted metatarsal plantarflexion,
a basic
requirement for 1st MTP joint dorsiflexion. With
this change, a planted
foot capable of permitting sagittal plane rotation up and over it was
born.
Hicks, in the Journal of Anatomy ('54) described how the dorsiflexing
hallux
caused the plantar aponeurosis to support the entire foot while
simultaneously creating an "irrestible" supinatory motion within the
foot.
In essence, he showed that while the body was permitted to advance
up and
over the foot, the foot could SIMULTANEOULSY become self stabilized
(a
principle later confirmed by Bosjen-Moller in 1979 in the J of Anatomy),
as
the forces maximized against it in the 2nd half of single support.
So, for bipedal gait to evolve, foot function became paramount.
Without a
structure which could permit sagittal plane advancement while also
self-stabilizing, upright gait would not be possible.
Looking forward to comments and further discussion.
Howard J. Dananberg, DPM
21 Eastman Avenue
Bedford, New Hampshire
USA
I am not a biomechanicist and only a beginner in
analyzing data from
kinetic perspective for gait. Therefore I don't intend to give the
discussion more insight about the energetics of human gait. I do want
to
look at gait at a dynamic theory point of view. This theory pointed
out
that all movement patterns were emerged as a cooperation of relevant
systems and the environment constraints, i.e. muscle strength, balance,
goal of walking, motivation, property of walking surface and level
of
crowdedness and etc.. The emerged pattern is the most stable pattern,
called an
attractor. It will take a higher level of energy in order to move it
out
of the attractor. Therefore, the energy required for walking should
be
the least, given all the current status of the relevant systems and
constraints. When one of the relevant systems is changed, i.e. being
in
a hurry or chased by a dog, the walking pattern will be no longer stable
but is pulled out of the current attactor until fall into another level
of attractor, i.e. fast walking or running. Thelen has successfully
explained gait development using dynamic theory. She also has
demonstrated that the disappearance of stepping reflex is due to the
fact that weight of the infant's leg outpaces his/her muscle strength,
nothing to do with the inhibition of higher level of nervous system
on
lower level as traditionally thought.
My point is if we compare
the energetics of different gait patterns
i.e. crawling (of babies) vs walking, walking in crowded place vs in
quiet place, walking without a goal vs with a goal, see what changes
most or least. That may give us more insight as to the dynamics of
human
gait, rather than only focusing on the gait itself.
I hope I am not off the theme of the discussion
too far.
Thanks for all the feedback on the evolution of gait. I thought I'd
better get my facts straight before I say any more, and I pulled out
my
copy of Origins by Richard Leakey this weekend - I now see just
what he
is getting at.
Darwin thought that bipedalism came as a package with intelligence and
tool-making, and this was later popularised by Owen Lovejoy,
who
emphasised the social consequences of the male being able to carry
food
back to the female. But fossil evidence doesn't bear this out.
If
Richard Leakey is correct (and I think he probably is), bipedalism
developed 7.5 mya (we still can't be sure because we still have no
human
fossils from so far back), which is a long time before any evidence
of
tools and language have been found (2.6 mya). Moreover, Lovejoy's
arguement depends on our ancestors being monogamous, which apparently
doesn't fit with the type of teeth and considerable body-size
dimorphism
of our ancestors.
He then makes the important point (for us) that in developing so early,
bipedalism must have been a simple practical solution to moving
around
the new grasslands whilst maintaining the ability to climb trees.
Many
people assumed that quadripedal gait (as studied in dogs or horses)
was
more efficient than bipedalism. Apparently, though, the energy
consumption of a chimpanzee (which is basically what humans were at
this
time) is the same when walking on 4 legs versus 2 (Rodman &
McHenry at
Davis - I'm afraid Leakey doesn't give the full reference). More
startlingly, human bipedal gait is considerably more efficient than
ape
knuckle-walking! So the transition from 4 legs to 2 seems to have been
a
quick pragmatic solution rather than a slow evolution. Of course, the
adaptations of the foot, knee and hip etc. would then have evolved
later, perhaps as bipedalism became more and more the norm.
So, "The" answer (assuming all this reasoning is correct - a lot of
"if"s, of course) would seem to be that we developed bipedal gait
because our previous method of locomotion (knuckle-walking) was a less
efficient method of moving around the newly forming grasslands in the
Rift valley. In other words, the apes basically made such a mess of
their locomotor apparatus by evolving it for climbing, that it became
ill-adapted for walking, so a new pattern (bipedalism) suddenly (shades
of dynamic pattern theory here, Louisa!) became advantageous.
Where does this leave us with respect to teasing out the underlying
principle of gait? Well, I've been looking again at the centre of mass
records of normal and pathological gaits, and what has struck me is
the
consistency of the motion in the one axis that Inman & Saunders
didn't
mention - the forward direction (a normal is at:
/teach-in/evolution/COM.avi
- the X
direction is the one I'm talking about). I'm amazed that even in
patients with quite severe walking disorders, the centre of mass
velocity seems to be almost constant. e.g the last Case of the Week:
/archives/16-7-99/CG.gif
I wonder whether our solution could be as simple as that: we walk
so as
to maintain forward velocity as constant as possible? Such a
pragmatic
goal would certainly fit with the above evolutionary arguement.
Look forward to your comments - I'd especially like to see any gait
recordings that do not show a constant CG velocity. I can't seem to
find
any in my own archives.
I would argue with the supposed date of the evolution of bipedal
gait as we
know it today as being 7.5ma. I know this is frequently quoted but
it is to
the best of my knowledge not based on anything substantial.
Huge assumptions are made which we really have no right the make. Much
of
the locomotor anatomy of the Austhralopithecus is far
from being adapted
for habitual bipedal gait, whatever the popular scientific press
will tell
us. The inominate bone of Afarensis is markedly different from modern
humans, perhaps being something of a hybrid (so to speak) between apes
and
humans. There is no complete Australopithicene feet - the one piece
that
could tell us most.
The nearest thing to this is the one foot of homo habilis
from 1.75ma - and
this has decidedly non-bipedal traits about it, in addition
to decidedly
bipedal traits - it is "missing link" stuff. The medial column is
ape-like
and the lateral column is human-like. However, the big assumption
we make,
which we have no right to do, is that it is anancestor of our. It could
simply have been an early experiment in bipedalism which went the way
of
the dinosaur....
Kind regards
Bob
Dr Bob Kidd BA Hons(Open) PhD(W.Aust)
Associate Professor
Head, Division of Podiatry
The University of Western Sydney - Macarthur
PO Box 555 Campbelltown
NSW 2560
http://FOHWEB.macarthur.uws.edu.au/podiatry/bob.htm
Phone: (02)46203633
Fax: (02)46203792
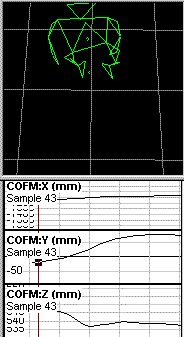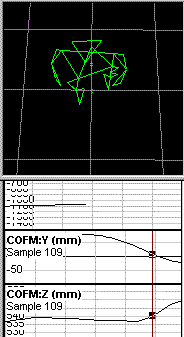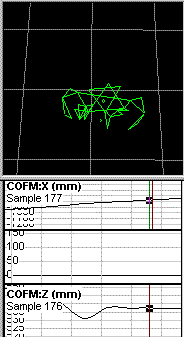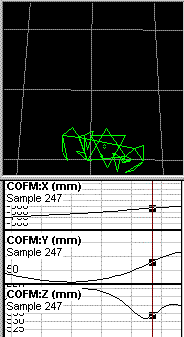
The plots that you refer to appear to represent position, or displacement,
of the center of mass, rather than instantaneous velocity of the
center of
mass. I am confident that if you calculate the first
derivative of the X
displacement, and expand it vertically to show the details,
you will find
that the line is not as straight as it appears to be in your plots.
When
displacement of the center of mass in the direction of progression
is
plotted at a scale which allows the total displacement to be observed,
the
variations in velocity cannot be visually discerned. That does
not mean
that they are not there, however.
I argued a couple of days ago that the center of mass of a walking
biped
must speed up and slow down with every step, because the center
of support
does not stay directly under the center of mass, but moves from
in front of
the center of mass to behind it during every single-support phase,
and back
again during every double-support phase. In other words,
it is impossible
to walk at constant instantaneous velocity. Progressional
velocity maxima
occur during each double-support phase, and minima occur during each
single-support phase. In normal gait, the maxima are quite sharp,
while the
minima are much more rounded.
The longer the steps are, the more the center of mass must speed up
and slow
down, because the extreme support points are farther ahead of and behind
the
center of mass. Consequently, a normal person who is able to
walk freely
with long steps will speed up and slow down more than a disabled person
who
is obliged to walk more slowly, with very short steps.
In any analysis of gait, it is desirable to include a graph of instantaneous
progressional velocity as a sort of data control, because it
shows clearly
whether the subject was walking at a uniform average velocity during
the
period of data collection, by showing whether successive maxima are
at a
uniform value and successive minima are at a (lower) uniform value.
Most of
our analyses presume that average velocity is constant, but without
checking
there is no way to be confident the subject was not speeding up or
slowing
down during data collection.
I wonder whether our solution could be as simple as that: we walk
so as
to maintain forward velocity as constant as possible? Such a pragmatic
goal would certainly fit with the above evolutionary arguement.
...
I don't think it can be that simple.
Someone once suggested (I don't remember who) that a rolling wheel
or barrel
with an off-center center of mass would demonstrate a path of
the center of
mass similar to that of a walking human. As the wheel rolled
along, the
center of mass would move up and down, and it would speed up and slow
down,
with each revolution. The lowest velocity would occur when elevation
is
highest, and the highest velocity when elevation is lowest, in a manner
somewhat analogous to a simple pendulum. Is there any food for
thought in
this idea?
Larry Lamoreux, Ph.D., Gait Dimension, Lafayette, CA, USA
I take your point about the COM velocity - in fact as soon as I sent my last message the thought occurred to me too (honestly!). I have plotted the COM velocity at:
/teach-in/evolution/COMvel.avi
Now, while it is not, of course, a completely flat line, I still think it is not bad. Obviously, it would be impossible to walk at an absolutely constant velocity, but this subject (one of my young PT students - chosen at random, believe me!) has a peak-to-peak amplitude of +/- 0.1 m/s (the scale on the .avi movie is in millimeters/s), which is a pretty small percentage (8%) of the overall COM velocity (1.2 m/s). As a comparison, the vertical COM velocity (plotted on the same scale) is around twice this. So all in all, I still think the constant forward velocity idea is plausible - well, at least as good a hypothesis as minimising vertical displacement.
But, as ever, I cherish your thoughts!
BTW, to control these quantities, there would have to be some way for the body to measure them - anybody any thoughts on how the CNS could monitor the COM, and whether it does in practice?
BTW, to control these quantities, there would have to be some way
for
the body to measure them - anybody any thoughts on how the CNS could
monitor the COM, and whether it does in practice?
I've only been semi-following this discussion, but had a thought on
this:
Bill Warren at Brown has done some interesting work on visual control
of locomotion, and how the CNS uses optical flow patterns to regulate
balance during locomotion. I've listed the papers of his that I know of
below. Assuming the head and COM were moving at approximately the same
speed during overground locomotion, changes in COM velocity would be reflected
in changes the velocity of the optical flow field as well.
Just a thought,
Jon
Bardy, B.G., et al. (1996). “Motion Parallax is Used to Control Postural Sway During Walking.” Experimental Brain Research, 111 (2): 271-282.
Warren, W.H., et al. (1996). “Visual Control of Posture During Walking: Functional Specificity.” Journal of Experimental Psychology: Human Perception and Performance, 22 (4): 818-838.
Warren, W.H. and Kurtz, K.J. (1992). “The Role of Central and Peripheral Vision in Perceiving the Direction of Self-Motion.” Perception & Psychophysics, 51 (5): 443-454.
Warren, W.H., et al. (1986). “Visual Control of Step Length During Running Over Irregular Terrain.” Journal of Experimental Psychology: Human Perception and Performance, 12 (3): 259-266.
Jonathan
Dingwell, Ph.D.
Postdoctoral Research Associate
Rehabilitation Institute
of Chicago
345 East Superior, room
1401
Chicago, Illinois, 60611
Phone: (312)
908-8738
FAX:
(312) 908-2208
Web: http://manip.smpp.nwu.edu/dingwell/
Dave
Tucker CA, Ramirez J, Krebs DE, Riley PO. Center of gravity dynamic
stability in
normal and vestibulopathic gait. Gait & Posture, 1998;8:117-123
David E. Krebs, PhD, PT
Professor and Director, MGH Biomotion Laboratory
MGH Institute of Health Professions
101 Merrimac Street
Boston, MA 02114-4719
V: 617 726 8016
F: 617 726 8022
http://WWW.MGH.HARVARD.EDU/depts/biomotion/staff/krebs.htm
Chris,
.1 m/s is about right -- again see Tucker et al but since that's
10% of the average walking velocity AND the max is .1 larger and the
min
is .1 smaller, that 20% variability seems to us to be substantial!
Dave
Consider the previously hypothesized barrel with offset center of mass
rolling down a hill. The center of mass rises and falls, speeds
up and
slows down, all without a CNS.
There is an astonishing (to me) web page from Cornell University
that shows
a completely passive mechanical model of alternating bipedal walking.
No
control system, no actuators at the joints, but it walks neatly down
a
gentle slope. Have a look, at:
http://tam.cornell.edu/programs/humanpower/pdw.html
Larry Lamoreux
Lafayette, CA, USA
Many thanks for the further comments, and for alerting us to the web-page
from
Cornell (http://tam.cornell.edu/programs/humanpower/pdw.html),
showing their
Jung-Gomez-Ruina model, which, as you say, is impressive.
So, are you saying, then, that "The" answer concerning bipedal gait
is
simplicity of control?
Incidentally, I'd like to see the forward velocity plots of these machines!
Also, why, to keep up the evolutionary theme, do some birds (all
birds?) have
their knees round the opposite way (flexion forwards)? I couldn't
imagine a
Jung-Gomez-Ruina model working with a backward knee - what could possibly
be
the advantage? I notice that one German word for walking "spaziergehen"
means,
literally (according to my rudimentary Deutsch), "to go like a sparrow"!
Just a quick comment in response to Chris's question about bird knees:
The answer is that birds' knees don't bend backwards - their knees bend
forward, but are tucked out of sight underneath their wings.
The knee of a
chicken, it turns out, is actually very similar in structure to that
of a
human being. For more on the knees of birds and other animals,
have a look
at this very interesting article:
Dye, S. F. (1987) An evolutionary perspective of the knee. JBJS, 69-A, 976-983.
Stephen Piazza, PhD
Assistant Professor
Departments of Kinesiology and Mechanical and Nuclear Engineering
Center for Locomotion Studies, 29 Recreation Building
The Pennsylvania State University
University Park, PA 16802
In "Psychology and Life Zimbardo suggests that
bipedalism and
encephalisation (brain growth) are THE defining characteristics of
the species homo sapiens. It is often assumed that we started to
walk--like an infant--first crawling on all fours then gradually
learning to stand. In this model we naturally learn
to walk as
we grow more intelligent. Chimps and Gorillas don't walk upright
simply because they are not as clever or advanced as we are and can't
conceive the benefits.
However, the evolutionary evidence does not agree. Our ancestors
walked upright before the increase in brain capacity.
The earliest paleoanthropological evidence of bipedalism (which is
not based on "form vs function" and "common ancestor"
controversies)
are two sets of footprints in Laetoli valley near the equator
in
Tanzania. These footprints are 3.6 million year old.
Genetic data suggests we may have had a common ancestor with the
chimps some 6-8 million years ago. Signs of intellect (homo
sapiens
sapiens) are only found about ?100 000 years ago. Attempts to
find the missing link or intelligent ape, resulted in the Piltdown
man, a scientific fraud involving a human skull and orangutan
bones!
As scientists we tend to believe "common sense theories" such as
learning to walk upright is a "good evolutionary idea", or is "more
efficient". These are all encephalocentric models, manifestations of
the "Piltdown man" syndrome.
What of the bipedal models of evolution?
Dudley Morton in his book Human locomotion and body form
mentions
the theory of Broom and Dart (African antipodes). Evidence suggests
that our upright posture developed because we were once brachiating
apes; that is apes that swing from tree to tree using our arms.
(This
incidently explains why we still swing our arms when we walk). Our
upright posture never evolved from a stooped quadripedal posture.
Only once the brachiating upright posture became habitual, did our
feet adapt to the ground. Evidence for this is in the
compressed nature of the human pelvis compared to the elongated
pelvis of brachiating apes. With our arms freed from locomotion
tasks, the spare brain capacity could be employed elsewhere.
Craig Nevin
(Anatomical engineer from the antipodes)
University of Cape Town, South Africa
The Jung-Gomez-Ruina walking figure on the Cornell web site is
actually based
on the mathematical work of Tad McGeer, PhD who published this
description in
1993 in the Journal of Theortical Biology. He demonstrated how
muscles were
only necessary to start, stop and manage perturbation. Once in
motion, only
very minor (if any) muscular activity is required. One
of the interesting
aspects is also the interrelationship between forward limb contact
and
toe-off of the trailing side. McGeer showed how the "foot
slap" following
forward limb heel strike created a "0 cost" toe-off maneuver of
the trailing
limb.
The Jung-Gomez-Ruina model is very dependant on the rocker
foot which
permits the top to pass up and over it. Using a square foot for
this
structure would clearly have a negative impact on its ability to "walk
without muscle action". When the human foot fails to permit
sagittal plane
pivotal function, then muscle activity based on the McGeer principle
of
"perturbation assistance" will cause the "cost of function" to raise.
Having
feet which work in the efficient rocker mode would seem to be required
for
bipedal gait to effectively evolve.
Howard J. Dananberg, DPM
21 Eastman Avenue
Bedford, NH 03110
USA
As a tracker of humans (following their foot falls) the wonderment
of how we got
to this bi-pedal position is of great interest.
My theory is that it came about as an efficient method of feeding
on plants that
were good tasting and were very desirous - to the point of chasing
off or even
killing other ape branch competition. Most unique physical characteristics
that
we see in the animal world seem to revolve around their diets. Giraffe's
long
neck - whales baleen screens. Consider an early ape branch that favored
a
vegetative growth 1 to 2 meters off of the ground ... finding it more
effective
to stay on two legs and pick with hands ... moving from bush to bush
without
dropping down to all fours would certainly net them more food.
I recall a documentary film on PBS about a tribe from the mountains
of the
Philippines, during the 1934? Worlds Expo in Chicago. They had
set up a live
display of these tribes people within the expo grounds. I remember
seeing
pictures of their feet, very similar to a gorilla's (with a splayed
big toe,
perhaps opposable in ability) ... they habitually climbed trees for
their main
diet item ... a de-evolution? or just an evolution to effectively gather
their
own diet preference?
1) are you sure it is the knee joint (ie analogy and homology)?
Are you
sure it is not the subtalar joint with much modified tarsus as seen
in eg
kangaroos.
2) If it is the knee, the answer may lie in the lower limb rotation
that
occurs in all mammals about 6-8 weeks into uterine life. That
is, what was
at the back is now at the front etc.
Let us not forget, in this respect, that bird is a reptile, and
a good
example of polyphyletic evolution: they are both homeothermic
and bipedal.
Thus, both evolved twice!
My theory is that it came about as an efficient method of feeding
on plants that
were good tasting and were very desirous
I think one has to put this in the context of the day.
The Red sea is an embryonic ocean, about 6ma old. The
typical start to
crustal spreading (soon to become sea floor spreading) is an thing
called a
tripal junction. Spreading starts at a point source whcih very
quickly
changes to being three rifts meeting at that point. Simple geometry
tells
us that this cannot continue so one of the arms "fails" and ceases
to
spread. The failed arm of the Red Sea is in fact the East African
rift
valley - a hot bed of hominid evolution in the same time scale.
Several things occurred as a result of Red sea formation/failed arm
rift
valley, including notable climatic changes which will have brought
with
them consequent changes to plant life - trees diminishing, grasses
becoming
more predominent. Trees brought shelter from heat - the place is heating
up.
My thought (well not mine originally) is that that trees disapearing
removed a habitat - or at least brought about a new selection pressure.
The
heating up, especially at a time of encephalisation will have brought
about
selective pressures to move the brain to a cooler environs -
it is about
3-5' cooler 1-2 metres above the ground that it is on the ground.
Simple,
stand up! This leaves hands free for gathering and allows you to se
further
for prey etc.
I understand there is evidence from contemporary skulls that blood
flow to
the brain increased hugely during this time, indication a cooling
mechanism, among other things.
In the context of the day is the way to see it, I think.
The problem I have, is that one can easily account for a quadripedal
hominoid, and equally for a bipedal hominoid. It is the transition
period of
one to the other which is not so easy to account for. What one
can say with
some certainty is that the selective pressures of the day must have
been
huge - they were pushing the proverbial uphill and keeping it there!
Dr Bob Kidd BA Hons(Open) PhD(W.Aust)
Associate Professor
Head, Division of Podiatry
The University of Western Sydney - Macarthur
One interesting question
- at least from a motor control/balance point of
view is why do some humans stand quite comfortably on one leg to
the
(apparent) amazement of others. I've always been very
comfortable standing on
one leg with the other leg bent and the foot resting just above the
knee of
the weight bearing leg - most people look at this and say something
on the
lines of "How do you do that?" or "Isn't that uncomfortable?".
I actually find it very
comfortable and have been doing it as long as I can
remember - and every now and then I met other people who say "Hey,
I do that"
or "My, sister does that and I can't figure out why" etc... It
seems that
it's easy and natural for a small number of people but not for the
majority.
If there was some evolutionary
advantage to it then no doubt in 100-200k
years you'd find everyone standing on one leg and the CGA listserver
would be
busy with folks wondering why we all do this. So my view on bipedal
gait is
"it was dumb luck - evolution made me do it" - I'm very much enjoying
the
comments from everyone on this topic.
Regards,
Edmund Cramp,
Motion Lab Systems, Inc.
4326 Pine Park Drive, Baton Rouge, LA 70809 USA
+1 225 928-4248 (voice, 2 lines), +1 225 928-0261 (fax)
http://www.emgsrus.com
<http://www.emgsrus.com
It is by nature that all energy systems aspire to lower energy states.
Bipedal locomotion is an energy conserving model of locomotion
which
approaches that of the the wheel in simplicity. It approaches the physics
of
wheel locomotion with the COM oscillating as close to zero as possible
based
upon anatomic constraints. Perhaps the concept of ideal normalcy with
zero
oscillation in position and accelleration in the COM is indeed, or
as close as
we can get, to a uniform theory of gait. All variation could then be
associated with compensation or lack thereof. Deviation or compensation
for
less than adequate sagittal plane motion needed for forward progression,
as
elucidated by Dannenberg, lends a great deal of credance to this model.
Readers on this list might want to have a look at the following website:
http://biology.uindy.edu/Biol504/HUMANSTRATEGY/16transition.htm
All the best,
Bipedal locomotion is an energy conserving model of locomotion which
approaches that of the the wheel in simplicity. It approaches
the physics
of wheel locomotion with the COM oscillating as close to zero as
possible
based upon anatomic constraints. Perhaps the concept of ideal
normalcy with
zero oscillation in positon and accelleration in the COM is indeed,
or as
close as we can get, to a uniform theory of gait.
Physics says that we can have zero progressional acceleration only
by having
zero step length. If the step length is non-zero, there
will be alternating
frictional forces between the foot and the ground that force the center
of
mass to accelerate and decelerate cyclically. Physics decrees
that a good,
healthy, free-swinging gait with generous step lengths cannot occur
without
cyclic accelerations and decelerations of the center of mass.
The "concept
of ideal normalcy with zero oscillation in position and acceleration
in the
COM" seems at odds with this fact.
At each walking speed, an individual tends to choose a particular
combination of step length and cadence. As speed increases, both
step
length and cadence tend to increase. It seems evident that there
is an
optimizing principle at work here, but the exact nature of that
principle is
not obvious. Is it purely mechanical -- the optimal way
to use the bony
structure? I think not. Is it muscular -- the optimal way
to make use of
the characteristics of muscles? Perhaps.
Observing, and analyzing, the way that people change walking speed may
help
to identify the underlying principles that govern gait.
Larry Lamoreux
Lafayette, CA, USA
This topic refuses to die, doesn't it? Can't say I'm disappinted - it
would be nice to finally get somewhere with it this time!
Thinking about what Larry was saying, I plotted the CoM acceleration
together with the difference between CoM and CoP (the 'total' CoP
combined from three force platforms: i.e. CoP = CoP1*F1 + CoP2*F2 +
CoP3*F3)/(F1+F2+F3). I've put it at:
/teach-in/evolution/CoMCoP.gif
I got a good fit between these two variables, with a bit of fiddling
caused, I think, by a slight error in the co-alignment of the origins
of
the kinematic and force platform coordinate systems, which caused a
slight drift with time.
I ended up with the following equation:
CoM Acceleration = 9(CoM - CoP) + 2.7*t
I guess I shouldn't be surprised that the fit is so good - as Larry
points out, this is straightforward physics. It's interesting, though,
that this is, of course, the same equation that Gurfinkel (and
later
Winter) used to characterise postural sway.
I wonder what the significance of the gain factor (9) is? Should
we
expect this to be the same for everyone, should it be dependant on
walking speed, stride length, or what?
Let me have your hypotheses while I analyse the requisite data...
Chris
--
Dr. Chris Kirtley
Dept. of Rehabilitation Sciences
The Hong Kong Polytechnic University
At each walking speed, an individual tends to choose a particular
combination of step length and cadence. As speed increases,
both step
length and cadence tend to increase. It seems evident that
there is an
optimizing principle at work here, but the exact nature of that
principle is
not obvious. Is it purely mechanical -- the optimal way to
use the bony
structure? I think not. Is it muscular -- the optimal
way to make use of
the characteristics of muscles? Perhaps.
If step length and cadence increase to increase speed, would that not
indicate that it is a mechanical change that is necessary to alter
speed. If
it was a muscular response, then the cadence or length would not need
to
change, only the power applied at each step.
Winter published a paper several years ago in which he showed muscular
action
in the weight bearing limb to be predominately eccentric during
single
support. Eccentric muscle contraction resists, not creates, motion.
Since
the speed of the CoM is increased during the 2nd half of single support
and
slowed by heel strike during double support, some mechanism other than
pure
muscle action (considering their eccentric nature of action) is necessary
to
create motion. The Jung-Gomez-Ruina walking model demonstrates
that muscles
are not needed for walking, but rather the intrinsic shape and function
of
the lower extremity appears to be able to walk by itself.
The eccentric
contraction, which is far more efficient than a concentric contraction,
could
maintain support to the weight bearing limb while the body raises and
lowers
its CoM as it steps up and over the weight bearing foot. A unifying
the
theory of gait will need take this into consideration.
Regards,
Howard J. Dananberg, DPM
Bedford, NH
USA
howiedbpg@aol.com
1. Roads (especially paved) are a relatively new creation.
Wheeled
structures with roads are quite efficient; take away roads and they
are less
so, and add an uneven (hilly) terrain and vegetation and suddenly legged
structures start looking pretty good. Especially those with at
least 4
legs, where postural stability is less of a challenge and natural rythmic
control/coordination schemes emerge. With bipeds, perhaps
posture and
balance are (nearly) everything -- just ask the wealth of roboticists
and
FES researchers, most of whom have been pretty humbled when trying
to
control biped locomotion. Andy Ruina's group and FES researchers
tell us
that the swinging through is pretty easy and potentially mechanically
efficient; yet gait is remarkably "inefficient" in that muscles
cocontract
and are often involved in eccentric contraction. It's
as if walking is an
aside to the challenge of just moving around WITHOUT FALLING
-- moving
around in the kitchen (of today or 2000 years ago), setting the table
or
making the bed, going over the hill to find the kids or watch the sunset.
It's also nice to watch the kids or sunset while moving -- so it's
best if
the head not bop up and down too much, so that the vestibular apparatus
isn't too stimulated and one can easily use the VOR and smooth pursuit
eye
movements to keep images stabilized. I doubt the locomotor
apparatus was
designed for gait -- I'd suggest it was designed to be robust
to a whole
collection of locomotor/postural tasks, with the "performance criteria"
certainly including a measure of efficiency at low effort, but first
and
formost related to safety from falling, and positioning effectly to
perform
everyday tasks.
2. Perhaps there's been too much emphasis on gait as a dynamic energetic
task?? Back in the 80's I had a grad student who was doing
3-D inverse
dynamics for various movement tasks, including gait, in older females
with
osteoarthritis. They moved so slowly that we decided to assume
behavior was
quasi-static and see if the results changed; to our surprise, when
we
overplotted the inverse dyamanics results with the inverse statics
(same
initial data), for about 95% of the gait task the two lines overplotted
such
that the difference was less less than the thickness of the lines!
And in
the few regions where they were distinct, they were close. So
we checked a
few young normals -- the lines still touched for a good deal of gait,
and
divergence was never more than about 10%. Add in the reality
that most
muscle activity seems more related to functioning as "stablizers",
perhaps
muscle impedance matters more than muscle force, and gait's
origins relate
first-and-foremost to stable performance for a rich variety of near-postural
tasks -- the movement as voluntarily-initiated perturbation
(temporary
controlled instability within the context of global stability) concept.
Then a few times a day the fast muscle fibers may need to kick in while
chasing something to eat (or a kid), or preventing a goal from being
scored).
3. Regarding insight into the GOVERNING PRINCIPLES of gait and a cybernetic
phenomena, I suggest more is learned through the scientific frustrations
of
dealing with physical objects and forward dynamic (and forward static)
simulations, versus inverse dynamics (where stability isn't a mathematical
issue). The inverse dynamic optimizations for gait
of the 1970's and 80's,
with the aims of figuring out "what was optimized" (e.g., energy,
force-squared, ...), were perhaps misleading in that maybe most of
the
muscles were working as
variable springs, and one of their functions was to be subservient
to
eye-head orientation (which in turn required a stable and relatively
smooth
COM). The forward optimization approaches of Pandy
and others are a great
direction, especially if their muscles include nonlinear spring-like
capabilities and there is an element of stochastic noise
(neural and/or
external perturbation) added in this system so that the predicted solution
has a margin of safety from falling.
Jack
PS - Larry L, nice to hear from you. The first gait analysis ever
performed
on me was in about 1981 in your lab at SF, when I was an early grad
student
at Berkeley -- I still have overheads of the data that you gave me,
and use
them in class!
{kind=link}
{kind=link}